Alfred Bernhard Nobel
- 化学賞
- 物理学賞
- 生理学医学賞
- 文学賞
- 平和賞
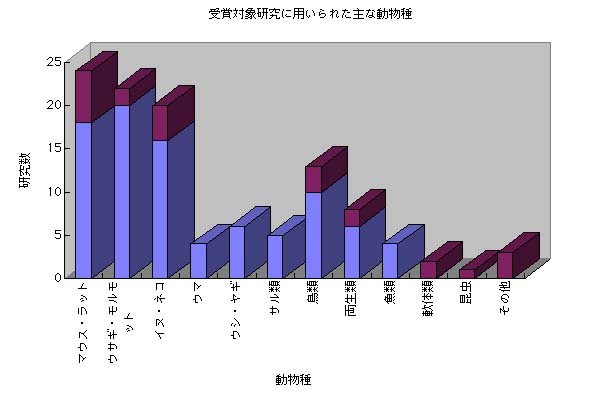
国別ノーベル生理学医学賞受賞数
(1901〜2025)

|
|
|
Alfred Bernhard Nobel |
ノーベル賞には5部門ある。
|
国別ノーベル生理学医学賞受賞数
(1901〜2025)
|
| 1901 | ベーリング(ドイツ) | 血清療法(特にジフテリアに対する研究)・・・モルモットで抗毒素発見 |
| 1902 | ロナルド・ロス(イギリス) | マラリアの侵入機構とその治療に関する研究(蚊の胃の中に原虫発見・ トリに感染) |
| 1903 | フィンセン(デンマーク) | 疾病の治療法への寄与ー特に光線治療法により医学の新領域開拓ー |
| 1904 | パブロフ(ロシア) | 消化に関する研究(イヌ) |
| 1905 | コッホ(ドイツ) | |
| 1906 |
ゴルジ(イタリア) カハール(スペイン) |
神経系の構造研究(ほ乳類、げっ歯類、鳥類) |
| 1907 | ラブラン(フランス) | 疾病発生における原虫類の役割に関する研究(患者赤血球の中にマラリア原虫の発見) |
| 1908 | エールリッヒ(ドイツ) メチニコフ(ロシア) |
免疫の研究 (1904 トリパンロート(色素剤)の発見・・・マウス) (1909梅毒治療薬サルバルサン606号発見・・・秦佐八郎 ウサギ、サル) (ミジンコ、ナマコ) |
| 1909 | コッハー(スイス) | 甲状腺の生理学、病理学および外科学的研究(コッヘル鉗子等も開発)(人体実験) |
| 1910 | コッセル(ドイツ) | 核酸物質を含む、タンパク質に関する研究による細胞化学の知見への寄与(魚卵) |
| 1911 | グルストランド(スウェーデン) | 眼の屈折機能に関する研究(人体実験) |
| 1912 | カレル(フランス) | 血管縫合および臓器の移植に関する研究(イヌの腎移植) |
| 1913 | リシェ(フランス) | アナフィラキシー・ショックに関する研究(イヌ) |
| 1914 | バーラーニ(オーストリア) | 内耳系の生理学および病理学に関する研究(人体実験) |
| 1915 〜 1918 | 第一次世界大戦中で受賞者なし | |
| 1919 | ボルデ(ベルギー) | 免疫に関する研究(動物の血清を調べる) |
| 1920 | クローグ(デンマーク) | 毛細血管運動に関する調整機構の発見(カエルなど) 比較動物学の草分け |
| 1921 | 受賞者なし | |
| 1922 | ヒル(イギリス) マイヤーホフ(ドイツ) |
筋肉中の熱発生に関する発見 筋肉の酸素消費と乳酸代謝の定式的関係の発見(カエル) |
| 1923 | バンティング(カナダ) マクラウド(イギリス) |
インシュリンの発見(イヌ) |
| 1924 | アイントホーフェン(オランダ) | 心電図の発見(生物の電気発生研究) |
| 1925 | 受賞者なし | |
| 1926 | フィビゲル(デンマーク) | 寄生虫発ガン説に関する研究(マウス) |
| 1927 | ワグナー-ヤウレック(オーストリア) | 麻痺性痴呆に対するマラリア接種の治療効果の発見(人体実験) |
| 1928 | ニコル(フランス) | チフスに関する研究(チンパンジー、モルモット) |
| 1929 | エイクマン(オランダ) ホプキンス(イギリス) |
抗神経炎ビタミンの発見(ニワトリ) 成長刺激ビタミンの発見(マウス) |
| 1930 | ラントシュタイナー(オーストリア) | ヒトの血液型の発見 |
| 1931 | ワールブルグ(ドイツ) | 呼吸酵素の特性および作用機構の発見(ウニ) |
| 1932 | シェリントン(イギリス) エイドリアン(イギリス) |
神経細胞の機能に関する発見(サル) |
| 1933 | モーガン(アメリカ) | 遺伝における染色体の役割に関する研究(ショウジョウバエ) |
| 1934 | ウイップル(アメリカ) マイノット(アメリカ) マーフィー(アメリカ) |
貧血に対する肝臓療法の発見(イヌ) |
| 1935 | シュペーマン(ドイツ) | 胚の成長における誘導作用の発見(イモリ) |
| 1936 | レーヴィ(アメリカ) デール(イギリス) |
神経刺激の化学的伝達に関する発見(カエル) (アセチルコリンの発見) |
| 1937 | ジェルジ(ハンガリー) | 生物学的燃焼過程、特にビタミンCおよびフマル酸の触媒作用に関する発見(ニワトリ) |
| 1938 | ハイマンス(ベルギー) | 呼吸調節における静脈洞と大動脈機構の役割の発見(イヌ) |
| 1939 | ドマーク(ドイツ) | (赤色染料)プロントジル(サルファ剤)の抗菌効果の発見(マウス、ウサギ) |
| 1940 〜 1942 | 第二次世界大戦中で受賞者なし | |
| 1943 | ダム(デンマーク) ドイジー(アメリカ) |
ビタミンKの発見 (ニワトリ) ビタミンKの化学的性質の発見(魚類) |
| 1944 | アーランガー(アメリカ) ガッサー(アメリカ) |
個々の神経繊維の高度に分化された機能に関する諸発見(ウシガエル) |
| 1945 | フレミング(イギリス) チェーン(イギリス) フローリー(オーストラリア) |
ペニシリンの発見、および種々の伝染病に対するその治療効果の発見 (ラット、マウス、ウサギなど多種) |
| 1946 | マーラー(アメリカ) | X線照射による突然変異体発生の発見(ショウジョウバエ) |
| 1947 | カール・コリ(アメリカ) ゲルティー・コリ(アメリカ) ウッセイ(アルゼンチン) |
グリコーゲンの触媒的分解経路の発見(動物の組織) 糖の物質代謝において脳下垂体前葉ホルモンの演ずる役割の発見 |
| 1948 | ミューラー(スイス) | 多数の節足動物に対するDDTの接触毒としての強力な作用の発見(サル) |
| 1949 | ヘス(スイス) モニス(ポルトガル) |
内臓の活動を統合する間脳の機能組織の発見(動物) ある種の精神病に対する前額部大脳神経切断の治癒的価値の発見(ロボトミー人体実験) |
| 1950 | ケンダル(アメリカ) ライヒスタイン(スイス) ヘンチ(アメリカ) |
諸種の副腎皮質ホルモンの発見およびその構造と生物学的作用の発見(イヌ、ブタ) |
| 1951 | タイラー (南アフリカ連邦) | 黄熱ワクチンの発明(蚊、サル、マウス) |
| 1952 | ワックスマン(アメリカ) | ストレプトマイシンの発見(動物、真核生物) |
| 1953 | クレブス
(イギリス) リップマン(アメリカ) |
クエンサン回路の発見(ほ乳類、鳥類) 補酵素Aと中間代謝におけるその重要性(動植物) |
| 1954 | エンダース(アメリカ) ウェーラー(アメリカ) ロビンス(アメリカ) |
種々の組織培地におけるポリオウィルス生育能の発見(サルや種々の実験動物) |
| 1955 | テオレル(スウェーデン) | 酸化酵素の性質及び作用機序の発見(馬の肝臓、酵母) |
| 1956 | クルナン(アメリカ) リチャーズ(アメリカ) ホルスマン(ドイツ) |
心臓カテーテルと循環器系の病理学的変化関する発見(イヌ) |
| 1957 | ボベット(イタリア) | クラレ様筋弛緩剤の合成に関する研究 |
| 1958 | ビードル(アメリカ) タータム(アメリカ) レダーバーグ(アメリカ) |
特定の化学現象を制御する遺伝子のふるまいの発見(1遺伝子1酵素説) (アカパンカビ) 細菌の遺伝的組換えおよび遺伝物質の組織化に関する発見(細菌) |
| 1959 | オチョア(アメリカ) コーンバーグ(アメリカ) |
リボ核酸およびデオキシリボ核酸の生合成機構の発見(大腸菌) |
| 1960 | バーネット(オーストラリア) メダワー(イギリス) |
後天的免疫寛容の発見 (動物実験によりバーネットの仮説を証明) |
| 1961 | ベーケーシ(アメリカ) | 内耳蝸牛における刺激の物理的機構の発見(ほ乳類) |
| 1962 | ワトソン(アメリカ) クリック(イギリス) ウイルキンズ(イギリス) |
核酸の分子構造および生体における情報伝達に対するその重要性の発見 (DNAラセン構造の解明) |
| 1963 | エクレス(オーストラリア) ホジキン(イギリス) ハクスリー(イギリス) |
神経細胞膜周縁および中心部の興奮・抑制にかかわるイオン機構の発見(イカ) |
| 1964 | ブロッホ(アメリカ) リュネン(ドイツ) |
コレステロールおよび脂肪酸代謝の機構と調節に関する発見(ラット) |
| 1965 | ジャコブ(フランス) モノー(フランス) ルヴォフ(フランス) |
酵素およびウイルス合成の遺伝的制御に関する発見(オペロン説)(大腸菌) (バクテリオファージとポリオウイルス) |
| 1966 | ラウス(アメリカ) ハギンズ(アメリカ) |
発がん性ウイルスの発見(ニワトリ) 前立腺がんのホルモン療法に関する発見 |
| 1967 | グラニト(スウェーデン) ハートライン(アメリカ) ワルド(アメリカ) |
視覚の化学的、生理学的基礎過程に関する発見 (節足動物、軟体動物、脊椎動物) |
| 1968 | ホーリー(アメリカ) コラナ(アメリカ) ニーレンバーグ(アメリカ) |
遺伝暗号とそのタンパク質合成における機能の解明 |
| 1969 | デルブリュック(アメリカ) ハーシー(アメリカ) ルリア(アメリカ) |
ウイルスの複製メカニズムと遺伝的構造に関する発見 |
| 1970 | カッツ
(イギリス) オイラー(スウェーデン) アクセルロッド(アメリカ) |
神経末梢部における液性伝達物質とその貯蔵、放出、不活化の仕組みに関する発見 |
| 1971 | サザランド(アメリカ) | ホルモンの作用機作に関する発見(cAMPに関する研究) |
| 1972 | エデルマン(アメリカ) ポーター(イギリス) |
抗体の化学構造に関する発見 |
| 1973 | ローレンツ(オーストリア) ティンバーゲン(イギリス) フリッシュ(オーストリア) | 動物行動学という学問分野を確立 (ミツバチ) |
| 1974 | デューブ(ベルギー) パラーデ(アメリカ) クラウデ(ベルギー) | 細胞の構造的・機能的構成に関する発見(小包体でのタンパク合成に関する研究)(ラット) |
| 1975 | テミン(アメリカ) ダルベッコ(アメリカ) ボルティモア(アメリカ) | 腫瘍ウイルスと遺伝子との相互作用に関する研究(マウス) |
| 1976 | ブランバーグ(アメリカ) | オーストラリア抗原の発見 |
| ガイジュセック(アメリカ) | 遅発性ウイルス感染症の研究(チンパンジー) | |
| 1977 | ギルマン(アメリカ) シャリー(ポーランド) ヤロー(アメリカ) | 脳内で産生されるペプチドホルモンに関する発見(ヒツジ) ラジオイムノアッセイ法の研究 |
| 1978 | アーバー(スイス) スミス(アメリカ) ネイサンズ(アメリカ) |
制限酵素の発見と分子遺伝学への応用(バクテリオファージ) (インフルエンザ菌) |
| 1979 | ハウンスフィールド(イギリス) コーマック(アメリカ) |
コンピューターを用いたX線断層撮影技術の開発(ウシの脳) |
| 1980 |
ベナセラフ(アメリカ) スネル(アメリカ) ドーセ(フランス) |
免疫反応を調節する細胞表面上の遺伝的構造に関する発見 |
| 1981 | スペリー(アメリカ) ウィーセル(スエーデン) ヒューベル(アメリカ) |
大脳半球の機能分化に関する研究 視覚系の情報処理に関する研究(ネコ) |
| 1982 | サミュエルソン(スエーデン) ベリストローム(スエーデン) ベーン(イギリス) |
プロスタグランジンの発見およびその研究 |
| 1983 | マクリントック(アメリカ) | トウモロコシを用いた染色体の研究でトランスポゾンの発見 |
| 1984 |
イェルネ(デンマーク) ケーラー(ドイツ) ミルスタイン(アルゼンチン) |
免疫制御機構に関する理論の確立とモノクローナル抗体の作成法の開発 (マウス、ウサギ) |
| 1985 | ブラウン(アメリカ) ゴールドスタイン(アメリカ) |
コレステロール代謝とその関与する疾患の研究(WHHLウサギ) |
| 1986 | モンタルチーニ(イタリア) コーエン(アメリカ) |
神経成長因子および上皮細胞成長因子の発見(ニワトリの胚) |
| 1987 | 利根川進(日本) | 抗体の多様性を生み出す遺伝的原理の発見(マウス) |
| 1988 | ブラック(イギリス) エリオン(アメリカ) ヒッチングス(アメリカ) |
薬物療法における重要な原理の発見 |
| 1989 |
バーマス(アメリカ) ビショップ(アメリカ) |
レトロウイルスのガン遺伝子が細胞起源である事の発見 |
| 1990 |
マレー(アメリカ) トーマス(アメリカ) |
人間の病気治療に関する臓器および細胞移植の研究(イヌ、ヒツジ、ブタ、ウシ) |
| 1991 |
ネーアー(ドイツ) ザクマン(ドイツ) |
細胞内に存在する単一イオンチャネルの機能に関する発見 |
| 1992 |
フィッシャー(スイス) クレーブス(アメリカ) |
生体制御機構としての可逆的タンパク質リン酸化の発見 |
| 1993 | ロバーツ(イギリス) シャープ(アメリカ) |
分断遺伝子の発見(人間を含む多くの生物) |
| 1994 | ギルマン(アメリカ) ロッドベル(アメリカ) |
Gタンパク質およびそれらの細胞内情報伝達における役割の発見 |
| 1995 | フォルハルト(ドイツ) ヴィーシャウス (アメリカ) ルイス(アメリカ) | 初期胚発生の遺伝的制御に関する発見(ショウジョウバエ、ゼブラフィッシュ) |
| 1996 | ドハーティー(オーストラリア) ツィンカーナーゲル(スイス) |
細胞性免疫防御の特異性に関する研究(マウス) |
| 1997 | プルシナー(アメリカ) | プリオン - 感染症の新たな生物学的原理 - の発見(ヒツジ、マウス、ハムスター、すべての動物) |
| 1998 | ファーチゴット(アメリカ) イグナロ(アメリカ) ムラド(アメリカ) |
循環器系における情報伝達物質としての一酸化窒素に関する発見(ウサギ、モルモット) |
| 1999 | ブローベル(アメリカ) | タンパク質が細胞内での輸送と局在化を司る信号を内在していることの発見 |
| 2000 | カールソン(スエーデン) グリーンガード(アメリカ) カンデル(アメリカ) |
神経系における情報伝達に関する発見 (アメフラシ) |
| 2001 | ハートウェル(アメリカ) ナース(イギリス) ハント(イギリス) |
細胞周期における主要な制御因子の発見(酵母) (ウニ) |
| 2002 | ブレナー(イギリス) ホロビッツ(アメリカ) サルストン(イギリス) |
器官発生とプログラム細胞死の遺伝制御に関する発見(線虫C・エレガンス) |
| 2003 | ラウターバー(アメリカ) マンスフィールド(イギリス) |
核磁気共鳴画像法に関する発見 |
| 2004 | アクセル(アメリカ) バック(アメリカ) |
におい受容体および嗅覚系組織の発見(ラット) |
| 2005 | マーシャル(オーストラリア) ウォレン(オーストラリア) |
ヘリコバクター・ピロリ菌の発見と胃炎および消化管潰瘍におけるその役割の解明(人体実験) |
| 2006 | ファイアー(アメリカ) メロー(アメリカ) |
RNA干渉-二重鎖RNAによる遺伝子サイレンシング-の発見(多くの生物) |
| 2007 | エヴァンズ(イギリス) カペッキ(アメリカ) スミティーズ(アメリカ) |
マウスの胚性幹細胞(ES細胞)を用いた、特定の遺伝子を改変する原理の発見(KOマウス) |
| 2008 | モンタニエ(フランス) バレ-シヌシ(フランス) ハウゼン(ドイツ) |
ヒト免疫不全ウイルスの発見 子宮頸癌を引き起こすヒトパピローマウイルスの発見 |
| 2009 | ブラックバーン(アメリカ) グライダー(アメリカ) ショスタク(イギリス) |
テロメアと テロメラーゼ酵素が染色体を保護する機序の発見(テトラヒメナ) |
| 2010 | ロバート・G・エドワーズ(イギリス) |
体外受精技術の開発 |
| 2011 | ブルース・ボイラー(アメリカ) ジュール・ホフマン(フランス) |
自然免疫の活性化に関する研究(マウス) |
| ラルフ・スタイマン(カナダ) | 樹状細胞と、獲得免疫におけるその役割の発見 | |
| 2012 | ジョン・ガードン(イギリス) 山中 伸弥(日本) |
成熟した細胞に対してリプログラミングにより多能性(分化万能性)を持たせられることの発見(マウス) |
| 2013 | ジェームズ・ロスマン(アメリカ) ランディー・シェックマン(アメリカ) トーマス・ズートホーフ(アメリカ) |
細胞内の主要な輸送システムである小胞輸送の制御機構の発見(イースト、ほ乳類の細胞) |
| 2014 | ジョン・オキーフ(アメリカ) マイブリット・モーセル(ノールウェイ) エドバルド・モーセル(ノールウェイ) |
脳内の空間認識システムを構成する細胞の発見(ほ乳類) |
| 2015 | ウイリアム・C・キャンベル(アイルランド) 大村 智(日本) チュ・ヨウヨウ(中国) |
線虫の寄生によって引き起こされる感染症に対する新たな治療法に関する発見(ほ乳類) マラリアに対する新たな治療法に関する発見 |
| 2016 | 大隅良典(日本) | オートファージの仕組みの解明(ほ乳類) |
| 2017 | ジェフリー・ホール(アメリカ) マイケル・ロスバッシュ(アメリカ) マイケル・ヤング(アメリカ) |
概日リズムを制御する分子メカニズムの発見 |
| 2018 | ジェームズ・P・アリソン(アメリカ) 本庶 佑(日本) |
免疫チェックポイント阻害因子の発見とがん治療への応用 |
| 2019 | ウイリアム・ケリン(アメリカ) |
細胞による酸素量の感知とその適応機序の解明 |
| 2020 | ハーベイ・オルター(アメリカ) マイケル・ホートン(イギリス) チャールズ・ライス(アメリカ) |
C型肝炎ウイルスの発見 |
| 2021 | デビッド・ジュリアス(アメリカ) アーデン・バタプティアン(アメリカ) |
温感と触感の受容体の発見(ほ乳類) |
| 2022 | スバンテ・ペーボ(スエーデン) | 絶滅したヒト族のゲノムと人類進化に関する発見 |
| 2023 | カリコ・カタリン(ハンガリー) ドリュー・ワイスマン(アメリカ) |
新型コロナウイルス感染症に対する効果的なmRNAワクチンの開発を可能にしたヌクレオシド塩基修飾(ほ乳類) |
| 2024 | ビクター・アンブロス(アメリカ) ゲリイリー・ラヴカン(アメリカ) |
マイクロRNAの発見と転写後遺伝子発現調節における役割の解明(C. elegans) |
| 2025 | メアリー・E・ブランコウ(アメリカ) フレッド・ラムズデル(アメリカ) 坂口志文(日本) |
抹消免疫寛容に関する発見 (制御性T細胞の発言を解明)(マウス) |